В конец
Глава 1. Атомы и молекулы
Атомы и молекулы. Тепловое движение частиц вещества.
Осмос
Осмос в жизни животных и растений
к оглавлению
в конец
Атомы и молекулы. Тепловое движение частиц вещества.
Как Вы уже, наверное, слышали, большинство окружающих нас предметов состоит из очень мелких, невидимых простым глазом частиц молекул. Молекулы, в свою очередь, состоят из еще более мелких частиц атомов. На рис. 1а молекулы воды, кислорода и еще нескольких веществ. На рис. 1б показано расположение молекул воды в жидкой воде, кристаллике льда и водяном паре.
Любые молекулы непрерывно двигаются (это движение называется тепловым движением). В жидкостях и газах молекулы летают по всей толще жидкости (по всему объему сосуда с газом), сталкиваясь и отскакивая друг от друга, как бильярдные шары. При повышении температуры жидкости (газа) они двигаются все быстрее и быстрее. Если понижать температуру, то скорость движения молекул будет уменьшаться, и наступит момент (при температуре 273° С), когда молекулы остановятся.
? Как, по-Вашему, должны двигаться молекулы твердых веществ? Почему Вы так думаете?
? На столе стоит стакан с водой. Что находится в пространстве между молекулами воды? Пожалуйста, выберите из следующих вариантов: вода, воздух, вакуум, стекло. Обсудите этот вопрос с учителем.
Любая группа молекул в пустоте (вакууме) самопроизвольно рассеивается (см. рис. 2: молекулам на периферии скопления ничто не может помешать улететь от него сколь угодно далеко). Если рядом оказывается несколько скоплений различных молекул, они, рассеиваясь, проникают друг в друга, и разные молекулы перемешиваются (пример растворение куска сахара в стакане с водой. Даже если воду не перемешивать, молекулы сахара за 23 дня рассеются по всему объему стакана, и раствор станет одинаково сладким в любой точке.) Любой подобный процесс называют диффузией.
Для описания диффузии удобно пользоваться словом "концентрация". Оно имеет несколько похожих, но все-таки разных значений. Вот самое простое из них. Концентрация это количество частиц данного вещества в единичном объеме (то есть в одном кубическом миллиметре, сантиметре или метре). Тогда диффузия - это процесс рассеивания вещества из области пространства с его высокой концентрацией в области пространства с его низкой концентрацией.
Осмос
Давайте проделаем такой опыт. Возьмем кусочек целлофановой пленки и привяжем его ниткой к стеклянной трубочке (рис. 3). Через эту трубочку наполним получившийся целлофановый мешочек сладкой водой (раствором обычного сахара). Теперь опустим трубочку с мешочком на конце в чистую воду так, чтобы уровни воды в трубке и в стакане сравнялись, и подождем некоторое время. Вода в трубке начнет подниматься все выше и выше над уровнем воды в стакане и, наконец, остановится.
Что произошло? Как Вы уже догадались, в целлофановой пленке имеются маленькие дырочки такого размера, что молекулы воды свободно через них проходят, а молекулы сахара "не пролезают" (рис. 4). Молекулы воды могут пролетать сквозь каждую дырочку как внутрь мешочка, так и наружу. Однако внутри мешочка часть объема занята молекулами сахара. Поэтому в непосредственной близости от дырочки внутри мешочка молекул воды всегда меньше, чем снаружи. Это значит, что число молекул, влетающих внутрь мешочка, всегда больше числа вылетающих из него, и постепенно в мешочке молекул воды становится все больше и больше. Вода заполняет мешочек и начинает вытесняться из него в стеклянную трубочку. Подобные явления называют осмосом.
Можно вспомнить о концентрации и сказать, что суть осмоса в том, что вода проникает сквозь целлофановую пленку из области своей высокой концентрации в область своей низкой концентрации, т.е. диффундирует. Иначе говоря, осмос это диффузия воды сквозь полупроницаемую перегородку (перегородку называют полупроницаемой, если сквозь нее не проходит хотя бы одно из разделенных ею веществ).
Осмос в жизни животных и растений
? Почему при варке варенья фрукты обычно сморщиваются? Попробуйте предложить способ приготовления варенья, при котором они бы не сморщивались.
? Почему у комнатных растений, которые забыли полить, листья повисают как тряпочки?
? Если вылить в чистую воду капельку крови, то имеющиеся в ней клетки начнут раздуваться и лопаться. Почему?
? Что произойдет с клетками крови в предыдущем опыте, если заменить чистую воду насыщенным раствором соли?
? Некоторые одноклеточные организмы (обыкновенная амеба, многие инфузории) обитают в почти чистой (пресной) воде. Почему же они не лопаются?
Глава 2. Строение клетки многоклеточного животного
! Что можно и чего нельзя увидеть простым глазом
! Царства органического мира
к оглавлению
в конец
Все живые существа Земли состоят из клеток маленьких мешочков размером обычно около 0,01 мм (мы будем пользоваться другой мерой длины - микрометром (общепринятое сокращение - мкм): 1 мкм = 1/1000 мм).
! Что можно и чего нельзя увидеть простым глазом
Есть такой способ оценки остроты зрения. Человеку предъявляют две черные точки на белом экране, а затем постепенно начинают их сближать (рис. 5). При нормальной остроте зрения и расстоянии от глаз до экрана 20-25 см эти две точки сливаются в одну при расстоянии между ними около 0,1 мм (то есть 100 мкм).
Реально объекты размером меньше 100 мкм на достаточно контрастном фоне (скажем, черный объект на белом фоне) видны как еле заметные точки, больше 100 мкм - видны с точностью до общей формы. Например, объект размером 100х200 мкм виден как маленькая палочка. На рис. 6 показан способ рассматривания мелких объектов, например, мелких организмов, обитающих в воде, зачерпнутой из пруда. Он позволяет различить (и, при желании, выловить пипеткой), например, протистов размером крупнее 50 мкм.
Оказывается, клетки разных организмов очень похожи друг на друга по строению. Если мысленно разрезать пополам какую-нибудь клетку многоклеточного животного, то на срезе будет видна картина, изображенная на рис.7.
! Царства органического мира
На Земле обитает несколько миллионов различных живых существ. Область биологии, называемая систематикой, занимается поисками наиболее удобных вариантов деления всего этого множества живых организмов на группы (они называются систематическими группами, или таксонами). Самые крупные таксоны - царства. Один из современных вариантов деления живых организмов на царства показан на рис. 8. В рамках этого учебника мы будем изучать только клетки многоклеточных животных. Клетки представителей других царств похожи на животные клетки, но все же заметно от них отличаются. На рис. 9 показаны для сравнения срезы клеток растений, грибов и бактерий. Царство бактерий имеет еще одно название - прокариоты ("про" "недо, еще не…", "карион" "ядро"), связанное с отсутствием в их клетках ядер. Остальные четыре царства носят общее название - эукариоты ("эу" - "настоящий").
Оболочку, покрывающую клетку снаружи, называют клеточной мембраной. Внутри клетки часто встречаются пузырьки, оболочка которых очень похожа на клеточную мембрану. Их называют мембранными пузырьками, или вакуолями. Различные части клетки называются органоидами. Видны срезы нескольких органоидов ядра, эндоплазматической сети (ЭПС), комплекса Гольджи, митохондрий, двух центриолей (вместе они имеют название "клеточный центр"). На рис. 6 показан внешний вид этих органоидов. Внутреннее содержимое клетки, за исключением ядра, называют цитоплазмой.
Клетка живет активной жизнью. Шевелится мембрана, разные органоиды перемещаются с места на место, некоторые мембранные пузырьки сливаются в один пузырек, другие, наоборот, разделяются на несколько новых пузырьков. На рис. 10 показаны различные "сюжеты из жизни" мембранных пузырьков. Если в роли большого пузырька на рис. 10б выступает вся клетка, то ситуацию выброса из клетки "начинки" мембранного пузырька называют экзоцитозом, а ситуацию захвата чеголибо внутрь клетки - эндоцитозом (от слов эндо и экзо).
Глава 3. Питание клетки
! Как клетка использует захваченные из внешней среды питательные вещества
Молекулярные механизмы фагоцитоза
Полимеры и мономеры
! Подробнее о полимерах, остатках мономеров и самих мономерах. Краткое введение в химию.
Строение мембраны. Липиды.
! Подробнее о липидах.
!Мембранное (пристеночное) пищеварение
к оглавлению
в конец
Любая живая клетка питается, т.е. захватывает из внешней среды съедобные для себя вещества (в виде отдельных молекул или больших групп молекул пищевых частиц, иногда даже целых клеток меньшего размера), и так или иначе использует эти вещества.
! Как клетка использует захваченные из внешней среды питательные вещества
Есть всего два принципиально различных варианта.
1. Молекулы питательных веществ можно использовать для построения других молекул, выполняющих в жизни клетки какиенибудь более или менее важные функции, например, различных молекул, входящих в состав клеточной мембраны. Этот вариант использования клеткой питательных веществ называется ассимиляцией.
2. Другой вариант - по сути дела сжечь их. Если поджечь, например, кусочек сахара или древесины, то он будет гореть, выделяя энергию в виде света и тепла. Клетка умеет производить похожий процесс (он называется дыханием) с отдельными молекулами пищи. Энергия, которая при этом выделяется, используется клеткой, например, для передвижения или для захвата новых пищевых частиц. Подробнее о дыхании мы поговорим в главе . Такой вариант использования веществ называется диссимиляцией.
Фагоцитоз (фагос пожиратель, цитос клетка) - питание клетки сравнительно большими пищевыми частицами (в том числе другими клетками). Общая картина фагоцитоза показана на рис. 11. Проплывающая мимо клетки пищевая частица касается мембраны и прилипает к ней. Мембрана под ней прогибается, охватывая частицу со всех сторон. В результате образуется мембранный пузырек с частицей внутри пищеварительная вакуоль. Она отрывается от мембраны и уплывает вглубь цитоплазмы. Там она сливается с другим пузырьком (первичной лизосомой - от слов лизис - растворение, расщепление и сома тело), отделившимся от комплекса Гольджи. Пузырек - результат этого слияния - называют вторичной лизосомой. После этого пищевая частица начинает растворяться. Минут через 20 внутри вторичной лизосомы виднеются только несколько маленьких бесформенных кусочков, почему-то "не захотевших" растворяться. Затем вторичная лизосома подплывает к мембране клетки и сливается с ней, выбрасывая из клетки наружу эти "кусочки".
Молекулярные механизмы фагоцитоза
Захват пищевой частицы (рис. 11: этапы 13)
Все эти удивительные превращения происходят благодаря деятельности специальных молекул. На рис. 12 а) показаны молекулы мембраны клетки (они называются рецепторами), обеспечивающие прилипание пищевой частицы к мембране и образование пищеварительной вакуоли. Рецепторы это молекулы мембраны клетки, которые могут узнавать другие молекулы лиганды, и прочно к ним прилипать. Коснувшаяся мембраны частица прилипает в том случае, если на ее поверхности имеются лиганды к какимнибудь рецепторам, имеющимся на поверхности клетки (на мембране обычно имеется около 100 различных разновидностей рецепторов, и каждый из них "узнает" определенный лиганд).
Растворение частиц пищи во вторичной лизосоме (рис. 11: этапы 57)
Пусть в данном конкретном случае клетка захватила с помощью фагоцитоза другую клетку, только маленькую. Первичная лизосома принесла из комплекса Гольджи специальные молекулы (пищеварительные ферменты), умеющие "разрезать" большие молекулы (например, полимеры - см. ниже) на части. Изза этого органоиды захваченной клетки "разваливаются" на отдельные мелкие молекулы (см. рис.12 б)). В мембране вторичной лизосомы имеются также белкипереносчики, которые умеют переносить эти мелкие молекулы через мембрану в цитоплазму клетки.
? Как Вы полагаете, откуда взялись в мембране лизосомы белкипереносчики?
Полимеры и мономеры
Молекулы состоят из еще более мелких частиц атомов. Полимеры ("поли" много, "мерос" часть) это молекулы, состоящие из одинаковых или очень похожих друг на друга групп атомов (остатков мономеров: "моно" - "один"), соединенных между собой (см. рис. 13 а)). Пищеварительные ферменты во вторичной лизосоме "разрезают" полимеры пойманной пищи на отдельные мономеры. Полимеры и их мономеры обычно имеют разные названия. Чтобы было легче запомнить эти названия, мы объединили в табл.1 сведения обо всех типах полимеров клетки.
! Подробнее о полимерах, остатках мономеров и самих мономерах. Краткое введение в химию.
Атомы в химии принято обозначать латинскими буквами (например, атом водорода обозначается латинской буквой H, атом кислорода буквой O). Молекулы в химии изображают формулами. В некоторых формулах (они называются графическими формулами) связи атомов друг с другом (химические связи) рисуют в виде палочек (см. например рис. 13 и 14). В аналитических формулах около значка каждого атома в виде индекса указывают количество таких атомов в этой молекуле. На рис. 14 показаны аналитические и графические формулы двух молекул - воды и перекиси водорода.
Обычно на одном конце любого клеточного полимера к нему присоединен атом водорода, а на другом конце - группа из двух соединенных друг с другом атомов - водорода и кислорода. На рис. 13 б) показано, как пищеварительный фермент "разрезает" полимер. Подобные химические реакции (в ходе которых к каким либо молекулам присоединяются разделенные на части молекулы воды) называют реакциями гидролиза. Пищеварительные ферменты, производящие реакции гидролиза, называют гидролазами.
Таблица 1. Полимеры и мономеры, входящие в состав живых клеток.
Полимеры |
Мономеры |
| Белки |
Аминокислоты
(обычно их в клетке около 20 разных типов).
|
| Углеводы (полисахариды): |
Моносахариды:
|
| Нуклеиновые кислоты: |
Нуклеотиды: |
| рибонуклеиновая кислота (РНК)
|
нуклеотиды РНК (4 типа: А аденин, У урацил, Г гуанин, Ц цитозин) |
| дезоксирибонуклеиновая кислота (ДНК)
|
нуклеотиды ДНК (4 типа: А, Т тимин, Г, Ц) |
Строение мембраны. Липиды.
Твердые предметы могут быть гидрофильными смачивающимися водой (филео - люблю) или гидрофобными не смачивающимися водой (фобос - страх). Определяется это так: на пластинку, изготовленную из интересующего нас вещества, капаем небольшую капельку воды. На гидрофобной поверхности капелька собирается в шарик, на гидрофильной растекается (рис. 15). Тонкая трубочка (капилляр) из гидрофильного вещества втягивает в себя воду. Капилляр из гидрофобного вещества вода как бы старается покинуть. Причина этих явлений в том, что молекулы воды притягиваются, во-первых, друг к другу (это притяжение мешает капельке воды мгновенно разлететься на молекулы), а во-вторых, к молекулам твердого тела. Если первая сила больше второй, то капелька воды собирается в шарик, "отлипая" от твердой поверхности (т.е. данная поверхность гидрофобная). Если первая сила меньше второй, то капелька воды начинает растекаться по твердой поверхности (т.е. данная поверхность гидрофильная).
Мембрана клетки состоит из многих разных молекул. Большинство из них молекулы липидов (рис. 16). Молекула липида состоит из гидрофильной "головки" и двух гидрофобных "хвостов". В воде "головки" разных липидных молекул слегка отталкиваются друг от друга, а "хвосты" слипаются друг с другом (вообще гидрофобные предметы в воде склонны слипаться друг с другом, при этом площадь соприкосновения с водой каждого из них уменьшается).
! Подробнее о липидах.
На рис. 17 а) изображены молекулы глицерина, фосфорной кислоты и двух разных жирных кислот. Если соединить их друг с другом так, как показано на рис. 17 б), получится липидная молекула, реально встречающаяся в мембранах некоторых клеток (подобные реакции называют реакциями дегидратации, то есть реакциями отделения воды). На рис. 18 показано еще несколько типов липидных молекул, встречающихся в мембране.
? Как Вы думаете, почему гидрофобные предметы в воде слипаются друг с другом?
? Подумайте, в каком положении будут находиться на поверхности воды одиночные липидные молекулы? А как будет выглядеть на поверхности воды пленка из молекул липидов толщиной в две молекулы?
В мембране, кроме липидов, имеется большое количество молекул белков (рис. 19). Одни из них погружены в двойной слой липидов (как вы думаете, какие участки их поверхности гидрофильны, а какие гидрофобны?), другие прикреплены к головкам липидов и целиком находятся либо снаружи, либо внутри клетки. Наружная поверхность клеточной мембраны отличается от внутренней: здесь к головкам липидов и к белкам прикреплены короткие цепочки из остатков моносахаридов олигосахариды. Все вместе они образуют на поверхности клетки "сахарную шубу" гликокаликс.
!Мембранное (пристеночное) пищеварение.
Некоторые белки могут довольно прочно прикрепляться к поверхности клетки, соприкасаясь при этом только с олигосахаридами гликокаликса. Например, такой способностью обладают многие пищеварительные ферменты. Если первичная лизосома сольется с наружной мембраной клетки, то многие из находившихся в ней пищеварительных ферментов, оказавшись "на улице", сразу же прилипнут к поверхности гликокаликса. При этом они смогут "ловить" проплывающие мимо соответствующие полимеры и расщеплять их. Белки-переносчики из лизосомы окажутся в наружной мембране, и начнут переносить внутрь клетки соответствующие мономеры. Получается, что пища будет перевариваться прямо на поверхности клетки. Особенно активно пристеночное пищеварение происходит в кишечнике у разных животных.
Глава 4. Полимеры клетки
Краткое введение в органическую химию
Строение молекул белков
! Моносахариды и полисахариды
Строение молекул нуклеиновых кислот (ДНК и РНК)
к оглавлению
в конец
Краткое введение в органическую химию
Органическими называют молекулы, в состав которых входят углерод и водород. Некоторые из них очень большие и сложные. Поэтому в органической химии (это наука, изучающая строение и свойства органических молекул) принято выделять внутри молекул характерные, часто встречающиеся группы атомов - функциональные группы. Каждая функциональная группа придает молекулам, в составе которых она встречается, определенные свойства. Поэтому удобно дать определенное название любым молекулам, в которых есть данная функциональная группа (см. табл. 2). Кроме углерода и водорода, в органических молекулах часто встречаются кислород (O), азот (N), сера (S), фосфор (P).
Таблица 2. Функциональные группы органических молекул.
Графическая формула функциональной группы |
Название функциональной группы |
Общее название всех органических молекул, содержащих такую группу |
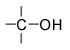 |
Гидроксилъная группа |
Спирты |
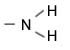 |
Аминогруппа |
Амины |
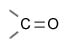 |
Кетонная группа |
Кетоны |
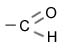 |
Карбонильная группа |
Альдегиды |
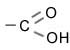 |
Карбоксильная группа |
Органические кислоты |
Строение молекул белков
Белки - полимеры, состоящие из аминокислот. Каждая аминокислота состоит из стандартной части молекулы (рис. 21) (она одинакова у 19 из 20 аминокислот), и вариабельной части (она у различных аминокислот разная). Две аминокислоты (только в пробирке: в клетке это всегда делается с помощью рибосомы) можно соединить друг с другом реакцией дегидратации. Два конца получившейся цепочки отличаются друг от друга по строению и свойствам , и называются N и C концы. Далее и к одному, и к другому концу можно присоединить новые аминокислоты, но в любом случае вся цепочка будет иметь с одной стороны N-конец, а с другой стороны C-конец. Такая нить из аминокислот называется первичной структурой белка 9 (рис. 22а)). Эта нить собирается в так называемые альфа-спираль либо в бета-структуру из-за взаимного притяжения стандартных частей аминокислот. Это вторичная структура белка (рис. 22б)). Среди вариабельных частей аминокислот данного белка есть гидрофильные и гидрофобные. Гидрофобные в воде слипаются друг с другом, вынуждая всю молекулу собираться в компактный комок (или глобулу, от английского слова globular - шарообразный), состоящий из нескольких соединенных между собой альфа-спиральных и бета-структурных участков. Это третичная структура белка (обычно она дополнительно укрепляется за счет образования химических связей между остатками аминокислоты цистеина, при этом получаются "мостики" из соединенных атомов серы - см. рис. 22в)). Наконец, иногда образуется сложная глобула, состоящая из нескольких слипшихся между собой глобул. Это четвертичная структура белка.
Если поместить молекулу белка в холодную воду, а затем начать ее медленно подогревать, то сначала (при самой низкой температуре, обычно около +50° С), разрушится четвертичная структура, затем - третичная, далее - вторичная, и, наконец, первичная.
? Белок куриного яйца и вправду представляет собой довольно концентрированный раствор различных белков. Попробуйте объяснить его превращение из жидкого в твердый в результате варки.
? (Очень трудный!) А почему белок при этом теряет прозрачность?
! Моносахариды и полисахариды
Моносахариды иногда еще называют альдегидоспиртами. В их состав входят карбонильные (альдегидные) и спиртовые группы (рис. 23а)). Существует номенклатура (то есть система названий) моносахаридов. Название любого моносахарида состоит из греческого числительного, означающего число атомов углерода в его молекуле (не менее трех!), и окончания "оза" (см. табл. 3). Все моносахариды могут самопроизвольно замыкаться в кольца, переходя из линейной формы в кольцевую (на рис. 23б) показан механизм этой реакции).
Таблица 3. Названия моносахаридов.
Число атомов углерода |
Греческое числительное |
Название моносахарида |
3
4
5
6
7
8
9
10 |
Три
Тетра
Пента
Гекса
Гепта
Окта
Нона
Дека |
Триоза
Тетроза
Пентоза
Гексоза
Гептоза
Октоза
Ноноза
Декоза |
Кольцевые формы моносахаридов могут вступать друг с другом в реакции дегидратации (рис. 24а)), образуя полимеры. На рис. 24б) - два разных полимера глюкозы (так называется одна из гексоз): крахмал и целлюлоза.
Строение молекул нуклеиновых кислот (ДНК и РНК)
Нуклеотиды состоят из соединенных вместе остатков фосфорной кислоты, какого-либо моносахарида (обычно рибозы или дезоксирибозы), и какого-либо азотистого основания (см. рис. 25 а) и табл. 4). Два нуклеотида (к сожалению, только в пробирке: в клетке это всегда делается совсем другим способом) можно соединить друг с другом реакцией дегидратации (см. рис. 25 б)). Два конца получившейся цепочки отличаются друг от друга по строению и свойствам, и называются 3' и 5' концы (читается "три штрих" и "пять штрих"). Далее и к одному, и к другому концу можно присоединить новые нуклеотиды, но в любом случае вся цепочка будет иметь с одной стороны 3'конец, а с другой стороны 5' конец.
Каждая молекула РНК (полное название рибонуклеиновая кислота) - полимер, состоящий из соединенных в цепочку один за другим остатков нуклеотидов РНК четырех типов (их обычно обозначают буквами: А - аденин, У - урацил, Г - гуанин, Ц - цитозин) (рис. 26 а)). Эта цепочка может изгибаться. Если при этом аденин случайно прикоснется к урацилу, то они немедленно прилипнут друг к другу. Точно так же слипнутся при встрече гуанин с цитозином. Принято говорить, что аденин комплементарен урацилу, а гуанин цитозину. Разумеется, соединение получится гораздо более прочным, если комплементарными друг другу окажутся не два случайно соприкоснувшихся нуклеотида, а целые группы (рис. 26 б)). Важно, чтобы прилипшие друг к другу комплементарные участки РНК были антипараллельными, то есть, чтобы направления от 3'- к 5'концу двух слипшихся участков были противоположными.
? Попробуйте придумать такую молекулу РНК, чтобы при слипании комплементарных участков друг с другом получилась:
1. "шпилька" (рис. 27 а)),
2. две соединенные друг с другом "шпильки" (рис. 27 б)),
3. "трилистник" (рис. 27 в)).
? Можно ли придумать такую молекулу РНК, чтобы при слипании комплементарных участков друг с другом получилась "петля" (рис. 28 г)? Почему?
ДНК похожа по строению на РНК, однако вместо урацила в ее состав входит другой нуклеотид тимин (он комплементарен аденину), и она состоит не из одной цепочки нуклеотидов, а из двух комплементарных друг другу антипараллельных цепей (рис. 29). Эти цепочки "скручены" в двойную спираль. Еще одно отличие: любой нуклеотид ДНК содержит моносахарид дезоксирибозу, а любой нуклеотид РНК моносахарид рибозу (см. табл. 4.)
Таблица 4. Сравнение ДНК и РНК.
Нуклеиновая к-та |
Моносахариды |
Азотистые основания |
| РНК |
рибоза |
А, У, Г, Ц |
| ДНК |
дезоксирибоза |
А, Т, Г, Ц |
Глава 5. Ассимиляция
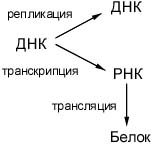
Сказка о передаче информации в клетке
Синтез новых белков (трансляция)
! Подробнее о реакции переноса цепочки аминокислот с одной молекулы тРНК на другую.
! Запуск (инициация) и завершение (терминация) синтеза белковой молекулы.
! Ядерные и цитоплазматические белки
Синтез новых молекул РНК (транскрипция)
! Созревание (сплайсинг и процессинг) РНК
Удвоение молекул ДНК (репликация)
! Присоединение нового нуклеотида к молекуле РНК или ДНК.
к оглавлению
в конец
Сказка о передаче информации в клетке
Представьте себе фабрику, выпускающую, ну, скажем, разнообразные кондитерские изделия. Много конвейеров, с одних один за другим сходят пирожки с повидлом, с других - торты "Птичье молоко", с третьих…….., и так далее. В центре завода - маленькая комната с большим числом дверей, а в ней - большая, можно сказать - огромная, поваренная книга. В комнату все время входят рабочие. Каждый такой рабочий некоторое время роется в книге, а затем выписывает из нее кулинарный рецепт-инструкцию, бросает его, снова роется в книге, выписывает следующий рецепт, и так далее. Готовые рецепты начинают беспорядочно плавать по комнате. Рано или поздно каждый рецепт выплывает наружу через дверь. Через несколько минут он доплывает до какого-нибудь конвейера, и этот конвейер начинает постепенно читать его, одновременно выполняя указанные в нем операции. Когда все операции выполнены, конвейер выгружает готовое блюдо, отпускает рецепт дальше плавать по фабрике, и захватывает какой-нибудь другой рецепт.
Время от времени оказывается, что фабрика достаточно разбогатела, чтобы приступить к созданию на ее основе двух дочерних предприятий. В какой-то момент в комнату начинают входить другие рабочие. Они постепенно переписывают книгу целиком, так что получается еще одна такая же книга. Однако в дверь книга не пролезает, поэтому перевозка одной из книг на новое место начинается с разборки комнаты на кирпичи. Затем из этих кирпичей заново собираются две новые комнаты. После этого все оборудование и персонал фабрики делится приблизительно пополам между двумя новыми фабриками.
В ролях:
| фабрика - клетка |
дверь - ядерная пора |
| конвейер - рибосома |
готовое блюдо - белок |
| комната - ядро клетки |
дочерние предприятия - дочерние клетки |
| книга - ДНК в ядре |
другие рабочие - много типов белковых молекул, участвующих в удвоении ДНК |
| рецепт - молекула иРНК |
кирпичи - мембранные пузырьки, на которые распадается ядерная оболочка |
Передача информации о структуре нуклеиновых кислот и белков:
Синтез новых белков (трансляция)
Синтез - это соединение нескольких молекул в одну, более сложную, молекулу. Синтез белка - сборка его молекулы из молекул аминокислот. Молекулы большинства белков собираются клеткой из 150300 аминокислот. Для того чтобы в правильном порядке соединить друг с другом эти аминокислоты, она должна иметь "инструкцию" для сборки каждого белка. В этой роли в клетке выступают молекулы информационной, или матричной, РНК. Причем одной аминокислоте соответствует группа из трех нуклеотидов - кодон.
?Сколько разных кодонов по три нуклеотида можно составить из четырех нуклеотидов РНК? Какое максимальное число аминокислот можно было бы закодировать, если бы каждый кодон состоял из двух нуклеотидов? Из одного нуклеотида?
Синтезом белков занимаются рибосомы - очень сложные молекулярные "машины". Каждая рибосома состоит из двух неравных частей. Их называют большая и малая субъединицы. Всего в обоих субъединицах одной рибосомы примерно 80 молекул белков и 4 разных молекулы РНК (РНК, входящую в состав рибосом, называют рибосомальной, сокращенно рРНК). Новые субъединицы рибосом образуются в ядрышке. Оттуда они выходят в цитоплазму через ядерные поры.
В цитоплазме обе субъединицы объединяются в целую рибосому на специальном стартовом участке любой молекулы информационной РНК (кратко иРНК или мРНК). Далее рибосома постепенно "проползает" всю молекулу иРНК до тех пор, пока не наткнется на сигнал прекращения синтеза белка один из кодонов УЦЦ, …………….. При этом из нее "вылезает" готовая цепочка соединенных друг с другом аминокислот. Дойдя до сигнала прекращения синтеза, рибосома "сваливается" с молекулы иРНК, при этом большая и малая субъединицы отделяются друг от друга и от готового белка (рис. 30).
Теперь подробнее о том, как это делается. Аминокислоты к работающей рибосоме приносят специальные молекулы транспортные РНК (кратко тРНК, см. рис. 31). Различные тРНК отличаются друг от друга антикодонами - тройками нуклеотидов, расположенными в определенном месте молекулы. В животных клетках обычно обнаруживается 31 тип тРНК, причем тРНК каждого типа занимается переноской одной определенной аминокислоты.
Специальные белки - кодазы (синоним аминоацил-тРНК-синтетазы) занимаются "подвешиванием" аминокислот к соответствующим тРНК. Каждый из них присоединяет к себе "свою" тРНК и "свою" аминокислоту (рис. 31), и затем соединяет их друг с другом.
? Как Вы думаете, сколько в клетке должно быть типов аминоацил-тРНК-синтетаз? Обсудите эту проблему с учителем.
В большой субъединице рибосомы есть два углубления А и Б, удобных для "посадки" молекул тРНК (рис. 32). Любая проплывающая мимо тРНК с висящей на ней аминокислотой может сесть в свободное углубление Б. Однако "прилипнет" она только в том случае, если ее антикодон окажется комплементарен кодону иРНК, в данный момент расположенному под этим углублением.
Когда это произойдет, вся цепочка аминокислот, висящая на тРНК в углублении А, будет прикреплена к аминокислоте, "приехавшей" на новой тРНК.
! Подробнее о реакции переноса цепочки аминокислот с одной молекулы тРНК на другую.
На рис. 33 показано, как это делается. По сути дела, между молекулами происходит обмен участками: атом водорода молекулы в А-участке рибосомы и цепочка аминокислотных остатков, висящая на тРНК в Б-участке, меняются местами.
тРНК из углубления А, лишившись аминокислоты, уплывает в цитоплазму. После этого рибосома делает "шаг" длиной в три нуклеотида как бы "мимо" тРНК, прилипшей своим антикодоном к иРНК. В результате эта тРНК с висящей на ней цепочкой аминокислот оказывается в углублении А. А в освободившееся углубление Б садится новая тРНК с аминокислотой, и все повторяется сначала. Цепочка аминокислот по мере выхода из рибосомы сворачивается во вторичную и третичную структуры.
! Запуск (инициация) и завершение (терминация) синтеза белковой молекулы.
Как уже рассказывалось, большие и малые субъединицы рибосом образуются в ядрышке. После выхода в цитоплазму малая субъединица соединяется с определенной молекулой тРНК (она называется инициаторная тРНК), на которой в этот момент уже "висит" аминокислота метионин (см. рис. 34). После этого малая субъединица присоединяет молекулы еще двух белков (они называются факторы инициации трансляции). К 5' концу иРНК присоединена специальная молекула - кэп (по-английски "cap" означает "шапочка, колпачок"). Малая субъединица, соединенная со всеми перечисленными молекулами, "узнает" кэп и начинает ползти от него по иРНК в сторону 3' конца. При этом антикодон инициаторной тРНК все время как бы ощупывает иРНК, и, в какой-то момент, прилипает к комплементарной ему тройке нуклеотидов - стартовому, или инициирующему, кодону АУГ. Малая субъединица останавливается и не трогается с места до тех пор, пока к ней не присоединится большая субъединица рибосомы. Затем к рибосоме присоединяется еще одна молекула тРНК, и начинается синтез белка. В начале любой белковой молекулы при таком способе синтеза, естественно, оказывается одна и та же аминокислота - метионин. Однако метионин впоследствии обычно "отрезается" специальным ферментом - гидролазой. Изготовление многих белков включает еще несколько операций (все вместе они обозначаются словами процессинг белков), например, вырезание участка из середины цепочки аминокислот, присоединение к вариабельной части какой-то аминокислоты остатка фосфорной кислоты и т.п.
Синтез белка всегда останавливается на одном из трех так называемых нонсенскодонов. Слово "нонсенс" означает "чепуха, бессмыслица". Дело в том, что эти три кодона не комплементарны никаким антикодонам тРНК. Поэтому рибосома, дойдя до нонсенскодона, останавливается (см. рис. 35). В цитоплазме имеются специальные белки, "вызволяющие" рибосому из подобного затруднительного положения. Они называются факторы терминации трансляции. Такой белок присоединяется к рибосоме, остановившейся перед нонсенскодоном, и гидролизует связь между тРНК и висящей на ее 3' конце готовой цепочкой аминокислотных остатков, то есть белком. В результате белок отделяется от рибосомы, и сама рибосома разваливается на большую и малую субъединицы.
Для полноты картины осталось рассказать, как попадают в комплекс Гольджи молекулы пищеварительных ферментов и белков-переносчиков. Синтез этих белков происходит на мембране ЭПС (рис. 36). Затем от ЭПС отделяются мембранные пузырьки с этими белками и сливаются с цистернами комплекса Гольджи. В комплексе Гольджи происходит сортировка всех доставленных белков. Результат этой сортировки - формирование вакуолей различного назначения с соответствующими молекулами внутри, например, первичных лизосом.
! Ядерные и цитоплазматические белки
Как видите, все белки клетки синтезируются в цитоплазме. Однако многие из них выполняют ту или иную работу внутри ядра. Как же они туда попадают, и что мешает оказаться в ядре белкам цитоплазмы? Ядерные белки имеют специальный "пропуск" - короткую цепочку определенных аминокислотных остатков. Белки, окружающие ядерную пору, захватывают белки с таким "пропуском", "заталкивают" их внутрь ядра, и больше уже не выпускают. Оказалось, что белки ядерных пор затягивают в ядро даже маленькие кусочки резины, если на их поверхности имеются описанные "пропуска".
Синтез новых молекул РНК (транскрипция)
В одной клетке в среднем около десяти тысяч разных белков, так что каждая клетка должна иметь целую "библиотеку" "инструкций по сборке" этих белков. Почему-то никакие из известных клеток не хранят полного набора молекул иРНК. Информация об аминокислотном составе белков хранится и передается по наследству от материнской клетки дочерним в виде молекул другой нуклеиновой кислоты ДНК. Гигантские (т.е. очень длинные) молекулы ДНК, хранятся в ядре клетки. По мере надобности специальный белок (РНК-полимераза) строит рядом с нужным участком ДНК молекулу РНК, комплементарную одной из двух цепочек ДНК (рис. 37). При этом РНК-полимераза узнает определенный участок ДНК - промотор, присоединяется сначала к нему, а затем начинает синтез РНК с точки, удаленной от промотора на определенное расстояние. Сразу после промотора обычно располагается оператор - участок, к которому могут прикрепляться различные регуляторные белки. Некоторые из них (репрессоры) мешают РНК-полимеразе прикрепляться к промотору. Другие (активаторы), наоборот, как бы делают промотор для РНК-полимеразы более липким. Далее располагается участок, РНКовую копию которого синтезирует РНК-полимераза (он называется "ген"), и, наконец, терминатор сигнал остановки транскрипции. Весь описанный участок ДНК, состоящий из промотора, оператора, гена и терминатора транскрипции, называется оперон.
Начиная транскрипцию, РНК-полимераза разъединяет две комплементарные цепочки ДНК на небольшом участке (рис. 38), синтезирует из нуклеотидов кусочек РНК, комплементарный одной из цепочек ДНК, а затем начинает двигаться, одновременно разъединяя перед собой цепи ДНК, присоединяя новые нуклеотиды к 3'концу кусочка РНК и вновь соединяя цепи ДНК позади себя. При этом за работающей РНК-полимеразой тянется "хвост" из свежеизготовленной РНК.
! Созревание (сплайсинг и процессинг) РНК
Любая молекула РНК до выхода из ядра наружу проходит сплайсинг, то есть специальные белки вырезают из нее ненужные участки (рис. 39). Это означает, что участок ДНК, с которого "считана" молекула РНК, содержал "бессмысленные" участки - интроны. "Осмысленные" участки ДНК, копии которых не вырезаются при сплайсинге РНК, называются экзонами. Про функции интронов еще далеко не все известно. Попробуйте предложить собственные идеи по этому поводу и обсудить их с Вашим учителем.
Многие типы РНК проходят еще и процессинг. Например, на 5'конец иРНК "навешивается" кэп, а на 3'конец - полимер, состоящий из приблизительно 200 остатков адениловой кислоты (его обычно кратко называют полиА).
Дальнейшая судьба новых молекул РНК может быть различной (см. рис. 40).
Новая молекула иРНК выходит из ядра в цитоплазму, там на нее оседают субъединицы рибосом и начинают синтез молекул закодированных в ней белков. Обычно в цитоплазме имеется небольшое количество пищеварительных ферментов - гидролаз, разрушающих молекулы РНК (рибонуклеаз). Они свободно перемещаются по цитоплазме, и каждая молекула иРНК рано или поздно встречается с таким ферментом, "разрезающим" ее на отдельные нуклеотиды. На этом деятельность этой молекулы иРНК заканчивается, и синтез на ней белков прекращается.
ДНК любой клетки имеет специальные участки, на которых точно так же синтезируются новые молекулы транспортных и рибосомальных РНК.
Синтез рибосомальных РНК в ядре первая стадия сборки целых субъединиц рибосом. В место, где это происходит (ядрышко), из цитоплазмы "приплывают" синтезированные там специальные ядерные "белки сопровождения", и налипают на молекулы рРНК, постепенно образуя будущие большие и малые субъединицы рибосом. В ядерной поре будущая субъединица проходит "переодевание": ядерные белки отделяются и уплывают вглубь ядра, так что в цитоплазму из ядерной поры выглядывают "голые" молекулы рРНК, и на них сразу же налипают "настоящие" рибосомальные белки (рис. 40).
Удвоение молекул ДНК (репликация)
В репликации (слово "реплика" - означает "отпечаток, копия") участвуют 5 различных белков (рис. 41). Все вместе они образуют так называемую репликативную вилку. Репликативная вилка постепенно ползет вдоль молекулы ДНК, оставляя позади две новые молекулы ДНК. Первой движется хеликаза. Она разъединяет две нуклеотидные цепочки ДНК. На образовавшиеся одноцепочечные участки немедленно налипают стабилизирующие белки. Стабилизирующие белки не дают двум комплементарным друг другу цепочкам ДНК вновь соединиться позади хеликазы. Следом за хеликазой по одной из цепей (она называется лидирующая цепь) ползет ДНК-полимераза в направлении к 5'концу. Она синтезирует новую цепочку нуклеотидов ДНК, комплементарную лидирующей цепи, присоединяя нуклеотиды ДНК к 3'концу. По второй цепи ДНК (отстающая цепь) ДНК-полимераза ползет в противоположном направлении (тоже в направлении к 5'концу). Но при этом получается, что отстающая цепь изготавливается "по кусочкам": ДНК-полимераза всякий раз ползет от хеликазы назад, к началу предыдущего кусочка, и отделяется от ДНК, оставив "дырку" между концом только что изготовленного кусочка и началом предыдущего. В эту "дырку" недостающие нуклеотиды вставляет специальный белок ДНК-лигаза.
! Присоединение нового нуклеотида к молекуле РНК или ДНК.
На рис. 42 показано, как это делается. Обратите внимание: в качестве "сырья" для изготовления нуклеиновых кислот используются не просто мономеры - нуклеотиды, а нуклеозиддифосфаты. Эти молекулы похожи на нуклеотиды, но, в отличие от них, содержат не один, а целых три остатка фосфорной кислоты. В результате каждой реакции присоединения нового нуклеотида (всегда к 3'концу!) "растущей" молекулы РНК или ДНК два фосфата отделяются.
? Попробуйте теперь заново прочитать "Сказку о передаче информации в клетке", и внести в нее все необходимые уточнения и исправления.
Глава 6. Цитоскелет.
Микротрубочки
! Механизм образования пищеварительной вакуоли при фагоцитозе.
Актомиозин.
Клеточный цикл. Митоз.
! Механизм деления цитоплазмы в клетках растений
к оглавлению
в конец
Любой из нас имеет скелет. Он состоит из твердых костей, гибких связок, соединяющих кости между собой, и мягких мышц, которые прикреплены к костям и, с силой меняя форму, изменяют взаимное расположение разных костей и мягких тканей тела относительно костей. В клетке имеются специальные белки, играющие роль костей и мышц. Всю систему таких белков называют цитоскелетом.
Микротрубочки
Микротрубочки (рис. 43) полностью соответствуют своему названию. Это прямые микроскопические трубочки (наружный диаметр 28 нм, внутренний 14 нм), состоящие из двух похожих друг на друга белков a-тубулина (a - греческая буква альфа, все слово читается "альфа-тубулин") и в-тубулина ("бета-тубулин"). Два конца микротрубочки отличаются друг от друга некоторыми важными свойствами (их называют "+" и "-" концы). В ДНК клетки имеются два разных гена, содержащие информацию о последовательностях аминокислот а-тубулина и в-тубулина. После синтеза на рибосомах в цитоплазме молекулы и тубулина объединяются в димеры (ди - два, мерос - часть). Димеры тубулина при определенных условиях могут присоединяться к "+"концу микротрубочки, микротрубочка при этом удлиняется. С "-"конца микротрубочки могут разбираться (то есть от него отделяются димеры тубулина, и микротрубочка при этом укорачивается). Изменяя условия в разных частях цитоплазмы, клетка имеет возможность делать сеть микротрубочек в ней более или, наоборот, менее густой. Кроме того, есть белки, способные присоединяться к "+"концам микротрубочек, прекращая тем самым их сборку, и другие белки, способные присоединяться к "-"концам и прекращать разборку микротрубочек.
Известны специальные транспортные белки, способные перетаскивать по микротрубочкам различные органоиды клетки. Один из них, кинезин, переносит их в направлении от "-" к "+"концу.
! Механизм образования пищеварительной вакуоли при фагоцитозе.
В большинстве клеток работают два независимых механизма.
Первый из них - простое следствие механизма прилипания пищевой частицы к мембране. За счет теплового движения молекул воды и пищевая частица, и рецепторы мембраны все время слегка вибрируют. Поэтому близко расположенные, но еще не соединившиеся друг с другом рецепторы и лиганды через короткое время сталкиваются и слипаются. Получается, что мембрана все больше и больше налипает на пищевую частицу со всех сторон (рис. 44а)).
Второй механизм обеспечивается работой специальных белков, одним концом присоединяющихся к рецепторам мембраны, уже прилипшим к лигандам на пищевой частице, а другим - к расположенным под мембраной микротрубочкам. Эти белки способны двигаться по микротрубочкам вглубь цитоплазмы, "волоча за собой" рецепторы, закрепленные в мембране. В результате работы многих таких белков весь кусок мембраны, прилипший к пищевой частице, погружается внутрь клетки, "на ходу" замыкаясь в пузырек (рис. 44б)).
Актомиозин.
Актомиозин - комплекс из молекул 4-х разных белков (а именно актина, тропонина, тропомиозина и миозина) в виде нитей в цитоплазме, способных с силой укорачиваться.
В результате синтеза белка на актиновой иРНК от рибосом отделяются молекулы G-актина (рис. 45а)). В цитоплазме они слипаются друг с другом в нити F-актина. Молекулы тропомиозина тоже сначала слипаются друг с другом в нити, а затем такая нить соединяется с нитью F-актина. На нить F-актина садятся также молекулы тропонина (рис. 45б)). Молекула тропонина состоит из трех субъединиц. Одна из них способна присоединяться к актину, вторая - к тропомиозину, а третья соединяет первые две, прикрепляясь одним концом к первой, а другим - ко второй. Нить, состоящую из этих трех белков, называют актиновым филаментом, или микрофиламентом.
Молекула миозина состоит из двух "головок" и "хвоста".Такие молекулы в цитоплазме могут слипаться друг с другом, образуя нити миозина (рис. 46). "Головки" молекул миозина образуют на поверхности нити миозина шесть продольных рядов. Отдельная молекула миозина в присутствии ионов кальция и АТФ перемещается по микрофиламенту в направлении от своего "хвоста". Нить миозина может присоединить максимум 12 актиновых филаментов (по 6 с каждого конца), и затем в присутствии ионов кальция и АТФ (подробно про ионы кальция рассказано в главе 7, а про АТФ в главе 9) "тащить" их друг к другу до соприкосновения (рис. 47а)). Выяснилось, что в некоторых клетках миозин образует димеры (рис. 47б)). Димер миозина может перемещать один микрофиламент по другому.
Клеточный цикл. Митоз.
Доказано, что новые живые клетки могут возникать одним единственным способом - в результате деления клеток. В ядре каждой клетки имеются молекулы ДНК, содержащие информацию об аминокислотном составе всех ее белков. Обе клетки, возникающие в результате деления, должны получить полноценные копии всех молекул ДНК материнской клетки. Для этого все молекулы ДНК материнской клетки должны быть сначала удвоены (период в жизни клетки, когда в ней происходит удвоение (репликация) ДНК, называется S фазой клеточного цикла), а во время деления клетки распределены по обеим дочерним клеткам.
Клеточный цикл - это последовательность событий, связанных с размножением клетки (рис. 48). Он состоит из собственно деления клетки (митоза), паузы до начала удвоения ДНК (G1 фаза), удвоения ДНК (S фаза) и паузы от момента окончания S фазы до начала митоза (G2 фазы). G1, S и G2 фазы вместе называются интерфазой.
Молекулы ДНК в G2 фазе перед началом митоза подвергаются тщательной упаковке с помощью специальных белков (рис. 49). Результат этой упаковки - митотическая хромосома. Перед началом митоза внутри ядра под микроскопом становятся видны хромосомы, соединенные попарно центромерами с помощью специальных белковых "замков" - кинетохоров. Каждая такая пара молекул ДНК - "сестры", получившиеся при удвоении одной молекулы ДНК клетки. При митозе им предстоит разойтись по разным дочерним клеткам.
Сам митоз состоит из четырех фаз: ПРОФАЗЫ, МЕТАФАЗЫ, АНАФАЗЫ и ТЕЛОФАЗЫ.
В профазе (рис. 50а)) происходит удвоение центриолей (каждая из двух центриолей материнской клетки строит около себя дочернюю центриоль) и две пары центриолей расходятся в разные концы (принято говорить: на разные полюса) делящейся клетки. После этого около каждой пары центриолей начинается сборка микротрубочек (при этом их "+"концы обращены от центриолей в цитоплазму). В результате образуется ВЕРЕТЕНО ДЕЛЕНИЯ, состоящее из двух половинок (ПОЛУВЕРЕТЕН) с парой центриолей в вершине каждой из них. В конце профазы оболочка ядра распадается на мелкие мембранные пузырьки (в конце митоза из них будут собраны два новых ядра), и хромосомы оказываются в цитоплазме.
В метафазе (рис. 50б)) "+"концы микротрубочек прикрепляются к кинетохорам хромосом. Первый из этих "+"концов может прикрепиться к кинетохору с любой стороны. Далее возможны два варианта развития событий. Если "+"конец второй микротрубочки прикрепится к кинетохору с той же стороны, что и первый, то в следующий момент кинетохор отделяется от обеих микротрубочек, и все начинается сначала. Если же "+"конец второй микротрубочки прикрепится к кинетохору со стороны другого полюса клетки, то кинетохор прочно прикрепляется к обеим микротрубочкам. Что происходит дальше, не вполне понятно. Почему-то сборка и разборка прикрепившихся к кинетохорам хромосом микротрубочек происходят так, что все хромосомы выстраиваются в плоскости экватора делящейся клетки. Известно, что если с помощью тонкой стеклянной иглы помешать одной паре хромосом добраться до этой плоскости, митоз приостановится до тех пор, пока эта пара не займет свое место.
Когда все хромосомы выстраиваются в экваториальной плоскости, специальные белки разрезают кинетохоры пополам, так, что "сестринские" хромосомы отделяются друг от друга и начинают расходиться к разным полюсам клетки. Это момент начала анафазы (рис. 50в)). Полуверетена в анафазе расходятся в разные стороны, причем каждое из них двигается как единое целое. Расхождение происходит за счет работы молекул белков, похожих на кинезин. Каждая такая молекула, прикрепившись к микротрубочке одного полуверетена, тащит ее по микротрубочке второго полуверетена в направлении к "+"концу (рис. 51).
В телофазе (рис. 50г)) происходит разборка микротрубочек веретена деления и образование двух ядер из мембранных пузырьков вокруг двух групп хромосом на полюсах клетки. Если стеклянной иглой отделить одну из хромосом от группы, то вокруг нее образуется отдельное маленькое ядро.
Последний этап митоза - деление цитоплазмы. У животных под мембраной клетки в районе ее экватора формируется кольцевой пучок актомиозина. Поочередно сокращаясь и перестраиваясь, он постепенно пережимает цитоплазму пополам, увлекая за собой мембрану.
! Механизм деления цитоплазмы в клетках растений
У растений экваториальная плоскость заполняется мембранными пузырьками, затем они сливаются друг с другом, разделяя цитоплазму на две части (рис. 52).
? Какие выводы можно сделать из опытов, описанных в рассказе про деление клетки? Предложите гипотезы:
1. о том, что мешает белкам, разрезающим кинетохоры хромосом, начать это делать до того, как все хромосомы окажутся в экваториальной плоскости клетки;
2. о том, что заставляет мембранные пузырьки в телофазе митоза собираться вокруг хромосом.
Глава 7. Векторные и скалярные величины. Сила.
Действия над векторами.
! Третий закон Ньютона.
к оглавлению
в конец
Векторными величинами, или векторами, называют величины, имеющие и численное значение, и направление. Например, если сказано, что автомобиль движется со скоростью 100 километров в час (то есть дано численное значение скорости), то про его скорость известно не все, потому что неизвестно, куда, в каком направлении он двигается. Примеры - скорость, сила, перемещение (перемещением движущейся точки в данный момент времени называют вектор с началом в точке начала ее движения, и концом в точке ее расположения в этот момент (рис. 53)).
Скалярными называют величины, имеющие численное значение, но не имеющие направления. Примеры - количество каких-нибудь предметов, длина, плотность.
Векторные величины обозначают в тексте буквами со стрелками (например,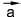 или
или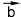 ), а на чертежах - стрелками, при этом длина стрелки равна численному значению вектора, а направление совпадает с направлением вектора (см. рис. 54).
), а на чертежах - стрелками, при этом длина стрелки равна численному значению вектора, а направление совпадает с направлением вектора (см. рис. 54).
Действия над векторами.
Действия над векторными величинами приходится производить с учетом их направления. В этом учебнике мы ограничимся рассказом о геометрических методах.
Сложение двух векторов (рис. 55а)). Для того чтобы сложить векторыи, нужно поместить начало векторав конец вектора. Тогда вектор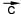 с началом в началеи концом в концеи будет равен их сумме. Точно так же можно складывать любое число векторов.
с началом в началеи концом в концеи будет равен их сумме. Точно так же можно складывать любое число векторов.
Умножение вектора на число (рис. 55б)). Вектор -представляет собой вектор с началом в концеи концом в начале. Вектор n представляет собой сумму n векторов.
Вычитание векторов (рис. 55в)). Вектор-можно представить как сумму двух векторов:-=+ (-).
? Пусть два человека тянут по полу рояль на двух веревках (рис. 56). Одна веревка действует на рояль с силой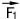 , другая с силой
, другая с силой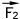 .
.
1. В каком направлении будет двигаться рояль?
2. С какой силой и в каком направлении нужно потянуть за третью веревку, прикрепленную к роялю в той же точке, что и первые две, чтобы он остановился?
! Третий закон Ньютона.
Взаимодействующие предметы действуют друг на друга силами одинаковой величины, направленными в противоположные стороны (см. рис. 57). Другая, более известная формулировка: "Действие равно противодействию".
Этот фундаментальный физический закон был сформулирован Исааком Ньютоном в году как обобщение наблюдений самых различных взаимодействий в природе, и с тех пор из него не обнаружено ни одного исключения.
В реальности этого закона легче всего убедиться, например, в невесомости, или на скользком льду, или на поверхности воды. Если в такой ситуации Вы оттолкнете от себя какой-нибудь достаточно тяжелый предмет, то сразу же полетите в противоположном направлении. Вы приложили некоторую силу к этому предмету, а он одновременно с этим - точно такую же силу к Вам, только направленную в противоположную сторону (рис. 57).
? Почему две противоположные силы, описанные выше, не компенсируют друг друга?
Глава 8. Электрические заряды. Основы химии.
Электризация предметов. Электрические заряды.
Строение атома
! Сила взаимодействия неподвижных зарядов. Закон Кулона.
Периодическая система элементов Д.И. Менделеева
Электроны в атоме
Электроотрицательность
! Электроотрицательность и закон Кулона
Химическая связь
! Ионные и молекулярные кристаллические решетки
Составление формул химических соединений
! Почему наружный электронный слой обязательно должен быть заполнен?
Вода. Растворы.
к оглавлению
в конец
Электризация предметов. Электрические заряды.
Давайте возьмем стеклянную палочку и будем 23 минуты с силой тереть ее кончик кусочком шелка. После такой процедуры палочка приобретает необычные свойства.
Во-первых, к ней начинают прилипать разнообразные мелкие предметы.
Во-вторых, она может передать это свойство другому предмету (для чего нужно потереть палочку об этот предмет).
В-третьих, самое удивительное, разные части этого последнего предмета начинают отталкиваться друг от друга (например, бумажные ленточки на рис. 57, отталкиваясь друг от друга, встают "дыбом"). Эбонитовая палочка, натертая шерстяной тканью, обладает точно такими же свойствами.
Однако бумажные ленточки, "заряженные" стеклянной палочкой, ПРИТЯГИВАЮТСЯ к бумажным ленточкам, "заряженным" эбонитовой палочкой.
Все описанные явления объясняются тем, что разные предметы при соприкосновении могут передавать друг другу мелкие, невидимые глазом, частицы - электроны. Они отталкиваются друг от друга, но притягиваются к другим частицам - протонам, которые имеются внутри любого предмета, но не могут переходить из одного твердого предмета в другой при соприкосновении. Протон отталкивается от другого протона, но притягивается к электрону. Свойство притягивать электроны и отталкивать протоны либо, наоборот, отталкивать электроны и притягивать протоны, физики называют электрическим зарядом. В первом случае заряд отрицательный (считается, что сам электрон тоже имеет отрицательный заряд), во втором положительный (считается, что сам протон тоже имеет положительный заряд).
? А теперь попробуйте объяснить все описанные выше события. Имейте в виду, что сила притяжения или отталкивания зависит от расстояния между частицами: чем ближе друг к другу находятся частицы, тем больше сила.
Строение атома
Любой атом состоит из ядра (оно находится в центре атома) и электронов, двигающихся вокруг него. Ядро обычно состоит из частиц двух типов: протонов и нейтронов (протоны, нейтроны, электроны и некоторые другие частицы, о которых в этой книге не рассказывается, называют элементарными частицами). Нейтроны не имеют электрического заряда, масса нейтрона почти равна массе протона. Масса электрона гораздо меньше: около 1/2000 массы протона. В любом атоме одинаковое количество протонов и электронов. Простейший атом (атом водорода) состоит из одного протона и одного электрона (рис. 57). Он не взаимодействует с заряженными предметами (т.е. не имеет электрического заряда) из-за того, что, например, к положительно заряженному предмету его электрон притягивается, а его протон отталкивается от этого предмета, причем эти две силы в точности равны друг другу. В подобной ситуации физики говорят, что два заряда различны по знаку, но равны по величине. Суммарный заряд любого атома равен нулю. Заряд электрона считают равным -1, а заряд протона +1.
Бывает, что один из электронов покидает атом. Число протонов в этом атоме оказывается на единицу больше числа электронов, а заряд его ядра на величину заряда одного электрона больше заряда всех электронов. Иначе говоря, суммарный заряд такого атома равен +1. Бывает, что атом захватывает извне добавочные электроны, приобретая суммарный отрицательный заряд. Атомы или группы атомов с добавочными или недостающими электронами называют ионами. Положительно заряженные ионы называют катионами, отрицательно заряженные анионами.
! Сила взаимодействия неподвижных зарядов. Закон Кулона.
Пусть имеются два неподвижных положительных заряда величиной +q1 и +q2 (то есть в первом из них протонов на q1 больше, чем электронов, а во втором - на q2), удаленных на расстояние R друг от друга (рис. .58). Первый из них действует на второй силой , второй на первый силой . По третьему закону Ньютона = . Будем теперь изменять q1, q2 и R, и посмотрим, как изменяется сила взаимодействия зарядов. Выяснится, что величина этой силы прямо пропорциональна величине каждого из зарядов (то есть увеличивается или уменьшается во столько же раз), а также величине 1/R2 (последнее означает, что при изменении R сила увеличивается или уменьшается во столько же раз, во сколько и 1/R2 , а не R; например, если R увеличилось в два раза, то сила уменьшится в 4 раза):
| |
|| ~ q1
|| ~ q2
|| ~ 1/R2
|
Эти три утверждения можно соединить в одно (мы не будем доказывать правильность такой замены, можете проверить ее на нескольких конкретных примерах):
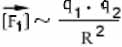
Однако эта формула еще не позволяет, зная заряды и расстояние, вычислить величину силы. Она позволяет только сказать, во сколько раз изменилась сила по сравнению с "начальной" ситуацией. Такой начальной ситуацией можно договориться считать вариант, когда оба заряда равны +1 (то есть заряду протона), а расстояние равно какой-то единице измерения расстояний, например, 1 метру. Предположим, мы действительно взяли два единичных заряда, поместили их на единичном расстоянии друг от друга, и измерили величину силы. Она оказалась равна k. Теперь можно сказать, что в нашей ситуации величина силы больше этого начального значения в (q1 · q2) /R2 раз:
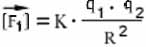
Это и есть математическая формулировка закона Кулона. Она скоро пригодится нам при обсуждении силы притяжения электронов к атомам.
? Ион имеет заряд +3, в нем 10 электронов. Определите заряд ядра.
Периодическая система элементов Д.И. Менделеева
Атомы сравнительно легко приобретают или теряют электроны, превращаясь в ионы. Гораздо труднее "заставить" атом потерять или приобрести протоны или нейтроны. В обычных условиях ядра большинства атомов никак не изменяются в течение сколь угодно долгого времени. Все известные науке варианты строения атомов объединены в таблицу, названную по имени ее создателя "Периодическая система элементов Д.И. Менделеева". Одну клетку в этой таблице "занимает" элемент, то есть все атомы Вселенной, имеющие одинаковый заряд ядра. Иначе говоря, все атомы данного элемента имеют одинаковое количество протонов, равное порядковому номеру этой клетки. Например, в природе существует три вида атомов с одним единственным протоном в ядре: протий вообще без нейтронов, дейтерий с одним, и тритий с двумя нейтронами (рис. 55). Любой из этих атомов является атомом водорода, и в таблице Менделеева находится в клетке N1. Атомы с одинаковым зарядом ядра, но различным числом нейтронов и, соответственно, с различной массой атома, называют изотопами ("изо" - "одинаковый", "топос" - "место"). Протий, дейтерий и тритий изотопы водорода.
Первые три строчки (их принято называть периодами) таблицы Менделеева изображены в табл. 6 (в каждой клеточке таблицы вверху - порядковый номер элемента, расположенного в этой клеточке, в центре - его обозначение (символ) в химических формулах, внизу - его название). Столбцы таблицы Менделеева называют группами.
Табл. 6. Первые три периода периодической системы элементов Д.И. Менделеева
группы
периоды |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
I |
1 H
водород |
|
|
|
|
|
|
2 He
гелий |
II |
3 Li
литий |
4 Be
бериллий |
5 B
бор |
6 C
углерод |
7 N
азот |
8 O
кислород |
9 F
фтор |
10 Ne
неон |
III |
11 Na
натрий |
12 Mg
магний |
13 Al
алюминий |
14 Si
кремний |
15 P
фосфор |
16 S
сера |
17 Cl
хлор |
18 Ar
аргон |
Электроны в атоме
Электроны в атоме непрерывно двигаются в пространстве около ядра. Есть физический закон (принцип неопределенности Гейзенберга), по которому задача точного прослеживания местонахождения движущегося электрона (в том числе - в атоме) в принципе не может быть решена. Можно лишь указать для каждого электрона область пространства внутри атома, внутри которой он находится столько-то процентов времени.
? Возьмите карту Вашего города и нарисуйте на ней область, внутри которой Вы находитесь:
а) не менее половины суток,
б) не менее 90 процентов суток.
Орбиталь - это область пространства, внутри которой данный электрон находится 90 процентов времени.
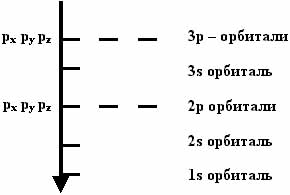
В атомах элементов I - III периодов встречаются s- и p- орбитали (см. рис. 56). В зависимости от расположения в пространстве различают px, py и pz орбитали; в зависимости от размера 1s, 2s и 3s; 2p и 3p орбитали (рис. 57).
Представьте себе ядро какого-нибудь атома (например, хлора), помещенное в вакуум, вместе с некоторым количеством свободных электронов. Один за другим эти электроны будут притягиваться к нему и "заполнять" орбитали вокруг этого ядра. Последовательность заполнения различных орбиталей вокруг ядра любого атома одинакова и показана в табл. 7.
Табл. 7. Порядок заполнения электронами орбиталей первых трех электронных слоев.
Заполнение электронами орбиталей данного атома происходит по следующим правилам:
1. на одной орбитали может располагаться не более двух (0, 1 или 2) электронов;
2. очередность заполнения: 1s, 2s, 2p, 3s, 3p. В этом списке каждая следующая орбиталь (или группа орбиталей, как в случаях 2p и 3p) заполняется только после того, как на каждой из предыдущих окажется по два электрона;
3. группы орбиталей, отличающихся только расположением в пространстве (например 2p и 3p), заполняются так, чтобы электроны располагались как можно "свободнее". Например, система 2р орбиталей заполняется в следующей последовательности (см. табл. 8, запись "2px1" означает, что на орбитали 2p, вытянутой вдоль выбранной нами координатной оси Ох, имеется один электрон).
Табл. 8. Порядок заполнения электронами системы 2р орбиталей.
| Общее число электронов на 2р орбиталях |
Расположение электронов на 2р орбиталях |
| 0 |
2px0 2py0 2pz0 |
| 1 |
2px1 2py0 2pz0 |
| 2 |
2px1 2py1 2pz0 |
| 3 |
2px1 2py1 2pz1 |
| 4 |
2px2 2py1 2pz1 |
| 5 |
2px2 2py2 2pz1 |
| 6 |
2px2 2py2 2pz2 |
Орбитали в атомах образуют электронные слои (1й слой: 1s, 2й слой: 2s и 2p, 3й слой: 3s и 3p). Орбитали одного слоя имеют одинаковый диаметр.
Электроотрицательность.
Электроотрицательность - это способность атома притягивать электроны наружного электронного слоя. Величина силы этого притяжения зависит, во-первых, от среднего расстояния от электрона до ядра (вместо этой величины можно использовать просто диаметр атома) и, во-вторых, от заряда внутренней части атома (атомного остова, то есть всего атома, за исключением наружного электронного слоя см. рис. 58). Диаметр атома постепенно уменьшается в пределах периода (увеличение заряда атомного остова "подтягивает" электроны ближе к ядру), и резко увеличивается при появлении нового электронного слоя, то есть в начале нового периода. Заряд атомного остова любого атома равен числу электронов наружного электронного слоя, взятому со знаком "+" (так как суммарный заряд атома равен нулю), и номеру группы в таблице Менделеева.
? Сравните электроотрицательность следующих атомов, не пользуясь таблицей Менделеева:
1. Li (№3) и B (№5),
2. Be (№4) и O (№8),
3. Mg (№12) и C (№6).
РЕШЕНИЕ п.3.
| Электронные формулы магния и углерода |
Mg: 1s2 2s2 2px2 2py2 2pz2 3s2 |
|
C: 1s2 2s2 2px1 2py1 2pz0 |
| Заряд атомного остова |
+2 |
< |
+4 |
| Радиус атома |
радиус Mg (3 эл. слоя) |
> |
радиус C (2 эл. слоя) |
Атом магния притягивает электроны наружного слоя слабее атома углерода по двум причинам:
1. наружные электроны магния располагаются в среднем дальше от ядра атома, чем наружные электроны углерода,
2. заряд атомного остова у магния меньше, чем у углерода.
ОТВЕТ: электроотрицательность Mg меньше электроотрицательности C.
! Электроотрицательность и закон Кулона
В только что рассмотренной задаче, по сути дела, был применен закон Кулона для сравнения сил притяжения электронов к двум разным атомам. Однако взаимодействие всего атома с одним из его электронов является взаимодействием не двух, а, например, для углерода, семи зарядов, шесть из которых (электроны) быстро двигаются. Поэтому перед началом расчетов строение реального атома приходится сильно упрощать (то есть использовать вместо самого атома его модель*). В явном виде список этих упрощений выглядит так:
1. Электроотрицательность не изменится, если заменить атомный остов точечным зарядом, расположенным в центре атома;
2. Электроны наружного электронного слоя располагаются в среднем на одинаковом расстоянии от ядра;
3. Электроны наружного электронного слоя в атоме не взаимодействуют между собой.
Иногда сравнить электроотрицательность двух атомов так, как это сделано в задаче, не удается (попробуйте сравнить электроотрицательности хлора (№17) и кислорода (№8)). Химики умеют измерять электроотрицательность атомов, проделывая для этого специальные эксперименты. Результаты этих измерений сведены в табл. 9 (в ней электроотрицательность фтора принимается равной 4,0).
Табл. 9. Электроотрицательность элементов первых трех периодов таблицы Менделеева
группы
периоды |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
I |
2,1 H
водород |
|
|
|
|
|
|
- He
гелий |
II |
0,97 Li
литий |
1,47 Be
бериллий |
2,01 B
бор |
2,50 C
углерод |
3,07 N
азот |
3,50 O
кислород |
4,10 F
фтор |
- Ne
неон |
III |
1,01 Na
натрий |
1,23 Mg
магний |
1,47 Al
алюминий |
1,74 Si
кремний |
2,10 P
фосфор |
2,60 S
сера |
2,63 Cl
хлор |
- Ar
аргон |
Химическая связь
Атомы в молекулах соединены между собой химическими связями. Они бывают ионными и ковалентными.
Ионная связь - это связь между положительным и отрицательным ионами. Самый простой способ образования ионной связи: при встрече двух разных атомов самый электроотрицательный из них отрывает от другого один или несколько электронов, превращаясь в анион. Второй атом, лишившись электронов, приобретает положительный заряд (то есть превращается в катион), и немедленно притягивается к отрицательно заряженному аниону (рис. 59).
Ковалентная связь возникает при взаимном проникновении друг в друга орбиталей двух атомов. При этом ядра обоих атомов притягиваются к области перекрывания их орбиталей (сильно упрощая реальную картину, можно сказать так. Представьте, что заряд электрона "размазан" по всему объему его орбитали. Тогда место, где перекрываются орбитали двух разных электронов, имеет "удвоенный" отрицательный заряд, и притягивает оба ядра см. рис. 60). Взаимное проникновение двух орбиталей с образованием связи возможно только в том случае, если на обоих этих орбиталях располагается в сумме не более двух электронов.
Ковалентная связь может быть полярной и неполярной.
Ковалентная связь неполярна, если связанные атомы имеют одинаковую электроотрицательность. В этом случае они с одинаковой силой притягивают зону перекрывания орбиталей, и в результате она располагается точно посередине между ними (рис. 60а)).
Ковалентная связь полярна, если связанные атомы имеют разную электроотрицательность. Тогда более электроотрицательный атом "подтягивает" зону перекрывания орбиталей ближе к себе (рис. 60б)).
Какая связь (ковалентная полярная либо ионная) возникнет между двумя разными атомами, зависит от того, насколько сильно отличаются их электроотрицательности.
! Ионные и молекулярные кристаллические решетки
Возьмем две молекулы фторида натрия NaF. Связь между натрием и фтором - ионная, заряды их +1 и -1. При встрече эти молекулы, разумеется, слипнутся в единую частицу (рис. ). Причем все четыре ионные связи в этой частице имеют одинаковую прочность. Если добавить еще несколько молекул NaF, они тоже прилипнут. Получившаяся система называется ионной кристаллической решеткой. Если разрушить ее, например, значительным повышением температуры, то вновь образуются отдельные молекулы NaF. Причем в одной молекуле могут оказаться "соседи" по кристаллической решетке, которые при формировании кристалла были в составе разных молекул. Говорят, что в ионной кристаллической решетке индивидуальность молекул не сохраняется.
Теперь проделаем те же операции с молекулами хлороводорода HCl. В этой молекуле ковалентная полярная связь. При встрече такие молекулы все равно слипнутся в единую частицу (рис. ), и к ней тоже прилипнут новые молекулы хлороводорода. Однако в получившейся молекулярной кристаллической решетке есть связи двух разных типов. Во-первых, это ковалентные полярные связи, "унаследованные" от отдельных молекул HCl. Во-вторых, это значительно более слабые связи между положительно и отрицательно заряженными концами молекул (без общих электронов, только силы Кулона). Если разрушить эту решетку повышением температуры, то образуются отдельные молекулы HCl. Причем в точности те же самые, что и при формировании кристалла. В молекулярной кристаллической решетке индивидуальность молекул сохраняется.
? Как Вы думаете, какие кристаллические решетки прочнее: ионные или молекулярные? Почему?
? Как связаны тип кристаллической решетки и температура плавления вещества? А температура плавления вещества и степень поляризованности ковалентных связей в его молекуле?
Составление формул химических соединений
Чтобы ясно представить себе сам процесс образования молекул из атомов, можно иметь в виду в качестве одного из возможных вариантов, например, такую картину. Большое число указанных в условии задачи атомов помещено в вакуум. В ходе теплового движения они сталкиваются и слипаются (то есть образуют химические связи) друг с другом. Готовой молекулой считается группа связанных между собой атомов, переставшая изменяться в результате этих столкновений.
? Известно, что молекула состоит только из атомов водорода и кислорода. Сколько атомов водорода и кислорода входит в ее состав, и как они связаны между собой?
РЕШЕНИЕ.
Мы разберем два способа решения подобных задач. В любом случае для начала выпишем электронные формулы обоих атомов.
H (№1): 1s1 O (№8): 1s2 2s2 2px2 2py1 2pz1
I способ.
Метод валентных связей.
Основа этого метода предположение, что все химические связи в будущей молекуле ковалентные. Еще одно правило: молекулы устроены так, что в них каждый атом имеет целиком заполненные орбитали наружного слоя (будем считать, что электроны, участвующие в образовании связи, принадлежат одновременно обоим связанным атомам).
Как видно из электронных формул, водород может образовать одну, а кислород не более двух связей (для заполнения внешнего слоя кислород должен образовать обе связи). Поэтому молекула состоит из одного атома кислорода и двух атомов водорода (рис. 61).
Теоретически решениями этой задачи являются также более длинные молекулы, содержащие неполярные ковалентные связи, например
Н-О-О-Н, Н-О-О-О-Н и так далее. Подобные молекулы действительно могут образоваться, однако при столкновениях с соответствующими атомами их неполярные связи легко разрушаются, заменяясь полярными ковалентными связями. Наиболее устойчивы цепочки из атомов углерода (с такими молекулами Вы уже знакомы) и кремния.
Число связей, образованных данным атомом в молекуле, называется валентностью этого атома. Обычно валентность можно предсказать с помощью метода валентных связей, однако иногда атомы могут проявлять неожиданные значения валентности. Например, строение молекулы угарного газа СО методом валентных связей предсказать нельзя. В подобных случаях используют более современные методы (например, метод молекулярных орбиталей).
II способ.
Метод ионных связей.
Метод основан на предположении, что все связи в молекуле ионные, то есть при образовании молекулы более электроотрицательные атомы отбирают электроны наружного слоя у менее электроотрицательных. Причем все ионы образующейся молекулы должны иметь заполненный наружный электронный слой.
Электроотрицательность кислорода больше электроотрицательности водорода, поэтому в рамках метода ионных связей можно считать, что атомы водорода отдают электроны атомам кислорода.
Степень окисления данного атома в молекуле это заряд данного атома, вычисленный исходя из предположения, что все химические связи этого атома ионные.
Каждый атом водорода в данном случае отдает свой единственный электрон (приобретая на внешнем слое заполненную 1s орбиталь и степень окисления +1). Каждый атом кислорода принимает два электрона (по одному на 2рx и 2рy орбитали), его степень окисления 2.
Число электронов, принятых атомами кислорода, и отданных атомами водорода, участвующими в образовании молекулы, должно быть одинаковым (физически это одни и те же электроны). В простейшем случае атом кислорода один, тогда атомов водорода - два.
ОТВЕТ: графическая формула Н-О-Н, аналитическая - Н2О.
! Почему наружный электронный слой обязательно должен быть заполнен?
Пусть мы наблюдаем за образованием молекул из отдельных атомов, сталкивающихся друг с другом и с частицами, получившимися при предыдущих столкновениях. Будем считать, что в данном конкретном случае образующиеся связи - ионные. В таком случае заданный вопрос сводится к трем более конкретным.
1. Может ли появиться молекула, в которой какой-то атом отдал атомам другого элемента не все электроны наружного слоя, и какова ее дальнейшая судьба?
2. Может ли какой-то атом, отдав атомам другого элемента при нескольких первых столкновениях все электроны наружного слоя, при очередном столкновении отдать хотя бы один электрон из следующего слоя?
3. Может ли какой-то атом, полностью заполнив наружный слой, присоединить еще хотя бы один электрон?
Давайте возьмем в качестве примера какой-нибудь атом, например, хлор, и посмотрим, как меняется его электроотрицательность по мере отнимания у него одного электрона за другим. Электронная формула: Cl (№17):
1s2 2s2 2px2 2py2 2pz2 3s2 3px2 3py2 3pz1
Заряд атомного остова +7, наружный электронный слой - третий. Используя нашу модель для сравнения электроотрицательности (см. стр. ), видим, что:
1. По мере отнимания семи наружных электронов заряд атомного остова и радиус атома не меняются, поэтому его электроотрицательность тоже не меняется. Ответ на первый вопрос: да, такая молекула может появиться, но при последующих соударениях оставшиеся электроны наружного слоя этого атома будут отбираться так же легко, как и предыдущие.
2. Когда семь наружных электронов отобраны, наружным слоем становится второй, а заряд атомного остова становится равным +15. В природе нет элемента, атомы которого обладали бы электроотрицательностью, позволяющей отнять электрон у такой частицы. Поэтому ответ на второй вопрос: безусловно нет.
3. Пусть хлор теперь принимает электроны. Если бы он принял 4s электрон, его наружным слоем стал бы 4s, а заряд атомного остова стал бы равным -2! Но в таком случае этот 4s электрон должен немедленно оттолкнуться от атома, ведь одноименные заряды отталкиваются! Поэтому ответ на третий вопрос - тоже нет.
Теперь попробуем предсказать строение молекул, состоящих из атомов азота и кислорода.
N (№7): 1s2 2s2 2px1 2py1 2pz1 O (№8): 1s2 2s2 2px2 2py1 2pz1
Электроотрицательность кислорода больше электроотрицательности азота, поэтому в рамках метода ионных связей можно считать, что атомы азота отдают электроны атомам кислорода
Азот в данном случае может отдать все пять электронов наружного слоя (при этом наружным становится заполненный первый слой, его степень окисления в этом случае +5). Кислород же принимает два электрона на 2р орбитали, его степень окисления -2: N+5, O-2.
Попробуйте сами придумать, как определить количество атомов каждого элемента в молекуле, если известны степени их окисления.
Надо заметить, что мы предсказали далеко не все существующие в природе варианты строения молекул, состоящих из азота и кислорода. Известны, например, молекулы состава NO, N2O , NO2, N2O3. Их строение можно предсказать с помощью метода молекулярных орбиталей.
Вода. Растворы.
Мы только что предсказали, пользуясь методом валентных связей, что водород и кислород образуют молекулу, состоящую из двух атомов водорода и одного атома кислорода. Это молекула воды. Обе ковалентные связи образованы 2р электронами кислорода, поэтому угол между ними должен составлять около 90 градусов. Сами связи - ковалентные полярные, так как кислород заметно превосходит водород по электроотрицательности. Это означает, что область перекрывания орбиталей смещена в сторону атома кислорода, следовательно, он имеет небольшой отрицательный, а атомы водорода небольшие положительные заряды. Тогда, во-первых, угол между связями в молекуле воды должен быть несколько больше 90 градусов из-за отталкивания друг от друга положительно заряженных атомов водорода (в действительности этот угол составляет примерно 108 градусов), во-вторых, молекулы воды должны стремиться повернуться друг к другу противоположно заряженными концами (рис.).
Связь между положительно заряженным атомом водорода из одной молекулы, и каким-либо отрицательно заряженным атомом из другой молекулы (например, кислородом), называют ВОДОРОДНОЙ СВЯЗЬЮ. Образование водородных связей снижает подвижность молекул воды, скрепляя их в группы (рис. ). Образованию таких групп препятствует тепловое движение молекул воды. Поэтому при понижении температуры возрастают и размеры таких групп, и процент молекул воды, принявших участие в образовании групп. Объем при этом, естественно, уменьшается. При замерзании все молекулы воды образуют друг с другом водородные связи (при такой температуре они успешно противостоят тепловому движению), формируя ажурную конструкцию (КРИСТАЛЛИЧЕСКУЮ РЕШЕТКУ льда) (см. рис. ). Видно, что упаковка молекул при этом оказывается менее плотной, чем в жидкой воде при 0° С, поэтому объем при замерзании воды увеличивается. Отдельные участки кристаллической решетки льда начинают образовываться уже при +4° С, поэтому при охлаждении воды объем начинает увеличиваться именно с +4° С.
Давайте теперь бросим в воду маленький кристаллик поваренной соли (формула NaCl). Кристаллическая решетка этого вещества состоит из катионов натрия и анионов хлора (см. рис. 1). Молекулы воды, естественно, "облепляют" находящиеся на поверхности кристалла ионы, поворачиваясь к ним противоположно заряженными концами, и, за счет теплового движения, соединенными усилиями отрывают их от кристалла. Оторванные ионы натрия и хлора плавают по всему объему раствора в "шубе" из прочно прикрепившихся к ним молекул воды.
ВОПРОС. Как Вы думаете, от каких особенностей поверхности твердого предмета зависит, будет она смачиваться водой или нет?
Глава 9. Энергия.
Полная энергия электрона в атоме.
Диссимиляция. Дыхание клетки
Переносчики атомов водорода
Дыхание
Гликолиз
Строение и работа митохондрий.
Цикл Кребса
Окислительное фосфорилирование
к оглавлению
в конец
Энергия - одно из самых важных физических понятий. Любой предмет (или, как говорят физики, любое ФИЗИЧЕСКОЕ ТЕЛО) обладает какой-то энергией. Различают три основных вида энергии: ПОТЕНЦИАЛЬНУЮ, КИНЕТИЧЕСКУЮ и ВНУТРЕННЮЮ.
Кинетической энергией обладает любое движущееся тело.
Кинетическая энергия тела массой m, движущегося со скоростью v, равна Е = Ѕ mv .
Тело обладает потенциальной энергией, если на него действует какая-то внешняя сила (например, оно притягивается к какому-нибудь телу, либо отталкивается от какого-нибудь тела). Например, потенциальная энергия тела массой m, поднятого над поверхностью земли на высоту h, равна E = mgh (здесь g множитель, учитывающий специфику планеты: для поверхности Луны g = , для Земли g = 9,81).
Внутренняя энергия тела это суммарная кинетическая и потенциальная энергия частиц (например, молекул или атомов), входящих в состав этого тела.
ПОЛНАЯ ЭНЕРГИЯ физического тела - это сумма его потенциальной, кинетической и внутренней энергий.
Для всех известных современной физике явлений справедлив
ЗАКОН СОХРАНЕНИЯ И ПРЕВРАЩЕНИЯ ЭНЕРГИИ:
Энергия не исчезает бесследно и не возникает из ничего, а может лишь переходить из одной формы в другую и от одного физического тела к другому.
Давайте обсудим конкретный пример полет камня, подброшенного с поверхности Земли вертикально вверх (рис. ). В момент броска человек, бросающий камень, придает ему начальную скорость V , то есть камень изменяет свою кинетическую энергию от нуля до величины Е = mV /2. Эту энергию он получает от руки человека, которая приводится в движение мышцами. Другими словами, кинетическая энергия сокращения мышцы (то есть движения ее концов навстречу друг другу) превращается в кинетическую энергию движения руки с зажатым в ней камнем. Затем камень отделяется от руки, унося с собой некоторую часть этой кинетической энергии. Рука после этого тормозится другими мышцами тела, при этом ее кинетическая энергия превращается в тепло (кто не верит, может вспомнить, как согревался в холодную погоду, "дрожа мелкой дрожью", то есть сокращая мышцы своего тела). Брошенный камень летит вверх, при этом его скорость уменьшается, а высота над землей увеличивается (то есть кинетическая энергия камня превращается в потенциальную). В верхней точке траектории кинетическая энергия равна нулю, а потенциальная максимальна. Затем начинается обратное превращение: высота уменьшается, скорость увеличивается. В момент возвращения в точку броска скорость камня равна скорости в момент броска, то есть его кинетическая энергия вернулась к начальному значению, а потенциальная энергия равна нулю. На самом деле последнее утверждения неточно: скорость (и полная энергия) камня немного уменьшилась из-за того, что летящий камень расталкивал молекулы воздуха, передавая им часть своей энергии. Как сказал бы физик, произошла потеря части энергии на ТРЕНИЕ о воздух.
ВОПРОС. Как Вы думаете, откуда взялась кинетическая энергия сокращений мышц? Может быть, она возникла из ничего?
Полная энергия электрона в атоме.
Свет.
Излучение и поглощение фотонов.
Полная энергия электрона в атоме не изменяется, пока он находится на своей орбитали. Поэтому для каждой орбитали любого атома можно указать точную величину полной энергии находящегося на ней электрона (рис. ). Промежуточные значения энергии для электрона "запрещены". Например, в атоме кислорода энергия электрона может соответствовать орбитали 1 или 2, но не может быть промежуточной между ними.
Электроны могут захватывать частицы света фотоны. В результате захвата фотона энергия электрона увеличивается, и он "перескакивает" на другую орбиталь, соответствующую новому значению его полной энергии. Разумеется, электрон может поглотить не любой фотон, а лишь такой, энергия которого в сумме с энергией этого электрона составляет "разрешенную" величину, то есть позволяет "перепрыгнуть" на определенную орбиталь, занятую в этот момент не более чем одним другим электроном. Совершив такой "прыжок", электрон недолгое время (порядка тысячной доли секунды) находится на новой орбитали, а затем "проваливается" обратно, излучая в случайном направлении точно такой же фотон, какой захватил раньше.
Диссимиляция. Дыхание клетки
АТФ - универсальный источник энергии в клетке.
Многие процессы, происходящие в клетке, требуют затрат энергии. Например, сокращение актомиозиновых нитей (именно оно лежит в основе укорочения мышц, переползания клеток с места на место, перемещения по клетке многих органоидов), перенос многих молекул сквозь мембрану, синтез белков и нуклеиновых кислот, транспортировка органоидов по микротрубочкам с помощью кинезина и т.д. Все белки, производящие в клетке подобные операции, одновременно расщепляют молекулы специального вещества - аккумулятора энергии АТФ (аденозинтрифосфорной кислоты) (см. рис. ). В этой молекуле связи между атомами фосфора и кислорода являются МАКРОЭРГИЧЕСКИМИ, то есть при их разрыве выделяется большое количество энергии. Белок, использующий АТФ как источник энергии, может разорвать две такие связи. При разрыве первой из них (с присоединением воды) получается молекула АДФ (аденозиндифосфорной кислоты) и фосфорной кислоты (рис. ). При разрыве второй макроэргической связи из молекулы АДФ получается АМФ (аденозинмонофосфорная кислота) и еще одна молекула фосфорной кислоты (рис. ).
Переносчики атомов водорода
Часто одним из этапов химического преобразования органических веществ внутри клетки является отделение от их молекул атомов водорода. Они присоединяются к специальным молекулам - переносчикам водорода, которые доставляют их в митохондрии на "сжигание". Обычно в клетке имеются три типа переносчиков водорода: НАД +, НАДФ + и ФАД. "Работают" они так (значок "" означает нековалентное присоединение):
НАД + + 2Н = НАД . Н + Н +
НАДФ + + 2Н = НАДФ . Н + Н +
ФАД + 2Н = ФАД . Н2
НАД + и НАДФ + захватывают один электрон (при этом атом водорода, у которого отобран этот электрон, превращается в протон и уплывает в раствор) и, кроме этого, нековалентно присоединяют еще один атом водорода. ФАД нековалентно присоединяет два атома водорода.
Дыхание
Как мы уже выяснили, клетка умеет производить процесс, аналогичный горению (он называется дыханием) с отдельными молекулами пищи. Энергия, которая при этом выделяется, используется клеткой для синтеза АТФ. В качестве сырья при этом используется только глюкоза (другие молекулы клетки для использования в качестве "горючего" сначала преобразуются в глюкозу) (здесь Фн - фосфорная кислота):
Как видно из схемы, в процессе дыхания одни и те же 6 атомов углерода, 12 атомов водорода и 18 атомов кислорода перешли из одного состояния (одна молекула глюкозы и 6 молекул водорода) в другое (6 молекул углекислого газа и 6 молекул воды). В результате выделилась энергия, поэтому, по закону сохранения энергии, суммарная внутренняя энергия этих атомов уменьшилась. На рис. сравнивается полная суммарная внутренняя энергия одной молекулы глюкозы и шести молекул кислорода с внутренней энергией шести молекул воды и шести молекул углекислого газа. Разница и представляет собой ту энергию, которая при горении выделяется в виде тепла, а при дыхании обеспечивает синтез АТФ.
Дыхание представляет собой большое число последовательных реакций, в ходе которых энергия выделяется маленькими порциями. Весь процесс обычно делят на три этапа: гликолиз, цикл Кребса и окислительное фосфорилирование. Мы не будем подробно рассматривать все реакции , а ограничимся общими описаниями перечисленных этапов и составлением суммарных химических уравнений. Гликолиз происходит в цитоплазме клетки вне митохондрий без участия кислорода, цикл Кребса и окислительное фосфорилирование - внутри митохондрий, окислительное фосфорилирование - при обязательном участии кислорода.
Гликолиз
В ходе гликолиза молекула глюкозы превращается в два остатка уксусной кислоты, которые захватываются специальным переносчиком - коэнзимом А, который в результате этой реакции превращается в ацетилкоэнзим А, или, кратко, ацетил КоА (рис. ):
Многие организмы (их называют анаэробами), в том числе некоторые многоклеточные животные, не потребляют кислород, синтезируя АТФ исключительно за счет глюкозы. Митохондрий в их клетках нет. Накапливающиеся продукты гликолиза они тем или иным способом выбрасывают в внешнюю среду. Примеры: дрожжи, молочнокислые бактерии, многие паразиты кишечника (аскарида, ленточные черви).
Строение и работа митохондрий.
Митохондрии - основной источник АТФ в клетке. Митохондрия имеет две мембраны: наружную и внутреннюю. Наружная мембрана гладкая, а внутренняя "смята" в складки в форме гребней или трубочек (они называются "кристы", а содержимое митохондрии, окруженное ее внутренней мембраной - "матрикс митохондрии"). Внутри митохондрии имеется замкнутая в кольцо длинная молекула ДНК и весь аппарат синтеза белков, в том числе собственные митохондриальные рибосомы (они сильно отличаются от рибосом цитоплазмы клетки). Впрочем, некоторые белки митохондрия все же получает из цитоплазмы.
Цикл Кребса
Цикл Кребса - кольцевая последовательность реакций (рис. ). Будем считать началом цикла молекулу щавелево-уксусной кислоты. В первой же реакции на нее переносится с ацетилКоА остаток уксусной кислоты (он состоит из двух атомов углерода, трех водорода, и одного атома кислорода), в результате получается изолимонная кислота. В ходе остальных реакций цикла перечисленные атомы отделяются от изолимонной кислоты и получающихся из нее молекул органических кислот, и в последней реакции снова получается щавелевоуксусная кислота. Точнее говоря, в каждом цикле от изолимонной кислоты и получающихся из нее молекул отделяются составные части остатка уксусной кислоты, присоединенного два цикла назад.
Теперь легко написать суммарное химическое уравнение цикла Кребса (мы написали его для двух молекул ацетилКоА, так как следим за судьбой молекулы глюкозы):
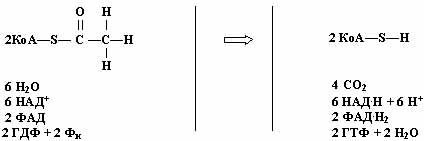
Окислительное фосфорилирование
Реакции цикла Кребса происходят в жидкости, заполняющей митохондрию, окислительное фосфорилирование - в ее внутренней мембране (рис. ) Здесь имеется пять типов белков дыхательной цепи, свободно перемещающихся в пределах этой мембраны (на рисунке они пронумерованы). Их функция - медленное поэтапное "сжигание" атомов водорода, доставляемых переносчиками НАД . Н и ФАД . Н2 с образованием молекул воды. Это делается так: белок №1 отбирает у НАД . Н электрон атома водорода и передает его белку №2, тот - белку №3, и так далее, до белка №5. Белки №2 и 4 имеют небольшие размеры, поэтому в мембране они двигаются значительно быстрее №№1, 3 и 5, и по сути дела, выполняют роль курьеров, разносящих электроны по назначению. При этом энергия электрона все время уменьшается (рис. ). Белок №5 накапливает четыре таких электрона, а затем производит реакцию образования воды:
4 е - + О2 + 4 Н - = 2 Н2О
Энергию, выделяющуюся при прохождении электрона по дыхательной цепи, белки №№ 1, 3 и 5 расходуют на выбрасывание протонов изнутри митохондрии в пространство между ее мембранами. В этом пространстве создается положительный заряд, а внутри митохондрии - отрицательный. Протоны, накопившиеся между мембранами, в этой ситуации имеют существенную потенциальную энергию за счет притяжения к внутренней части митохондрии. Во внутренней мембране, кроме белков дыхательной цепи, имеются молекулы еще одного белка - АТФ-синтетазы. Они пропускают протоны внутрь митохондрии, используя их потенциальную энергию для синтеза АТФ.
к оглавлению